被子植物的双受精是一个极为精细且高度特化的过程:两个精细胞分别与卵细胞和中央细胞融合,从而形成胚和胚乳。这一过程的核心在于雌雄配子的精准识别、黏附与融合。长期以来,配子识别与融合的分子机制是植物有性生殖领域的核心问题之一,该机制的解析对于理解植物有性生殖及其在农业上的应用具有重要的指导意义。在全球人口持续增长与粮食安全压力不断加剧的背景下,发展高效育种技术至关重要。与依赖多代自交的传统育种相比,双单倍体技术通过单倍体诱导和染色体加倍的方式,可以在两代内获得完全纯合的双单倍体品系,大大缩短育种周期。因此,鉴定高效和广泛适用的单倍体诱导基因对于加速作物改良具有重要意义。
近日,中国科学院遗传与发育生物学研究所李红菊研究组与杨维才研究组合作,在Plant Communications背靠背发表题为The sperm cell surface protein GEX3 acts with GEX2 and DMP8/9 to control gamete adhesion and fusion in Arabidopsis和Haploid induction by editing the conserved GEX3 gene in monocots and dicots的两篇研究论文,揭示了被子植物配子识别与融合的关键分子机制,并在单倍体诱导技术上取得重要突破。
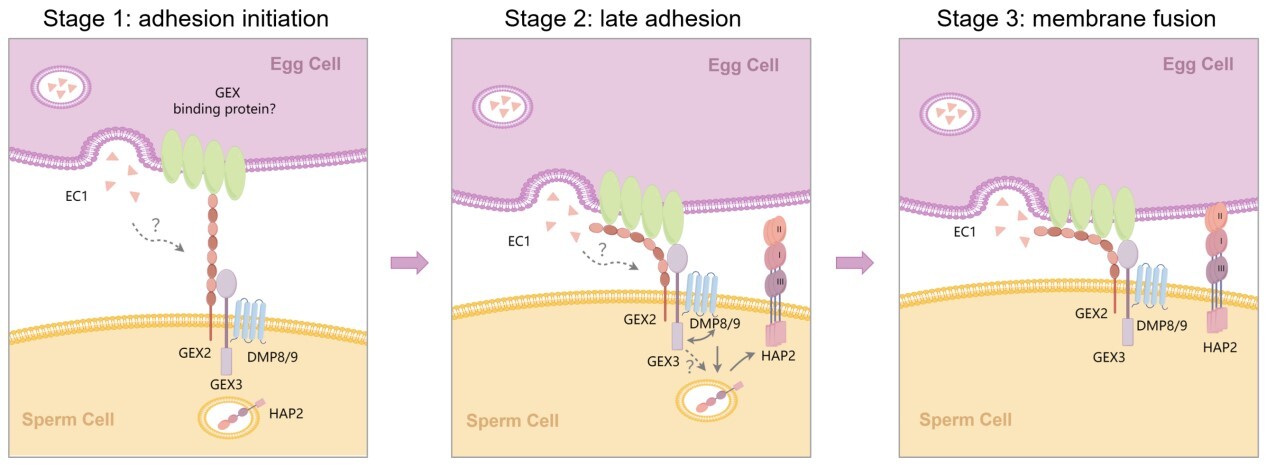
在第一项研究中,研究人员发现一个具有胞外β-螺旋桨结构域的精细胞表面蛋白GEX3是配子粘附和融合的关键因子。当GEX3突变后,突变体角果表现出严重的败育表型。gex3突变体的花粉活力、花粉管生长以及向胚珠的靶向均正常,但精细胞由于无法正常完成双受精而停滞在胚囊内。进一步分析显示,GEX3缺失既会造成配子粘附失败,也会造成配子融合失败,但对配子粘附的影响更大。为了解析其作用机制,研究人员通过多突变体分析和蛋白互作分析,证明GEX3与另一个配子粘附蛋白GEX2通过胞外域相互作用形成复合体,介导雌雄配子间的粘附。同时,GEX3与精细胞膜蛋白DMP8/9协同作用促进融合蛋白HAP2从胞内转移至精细胞质膜,以确保配子质膜融合。在此基础上研究人员提出了双受精分子调控的三阶段模型。
GEX3介导配子粘附和融合的工作模型
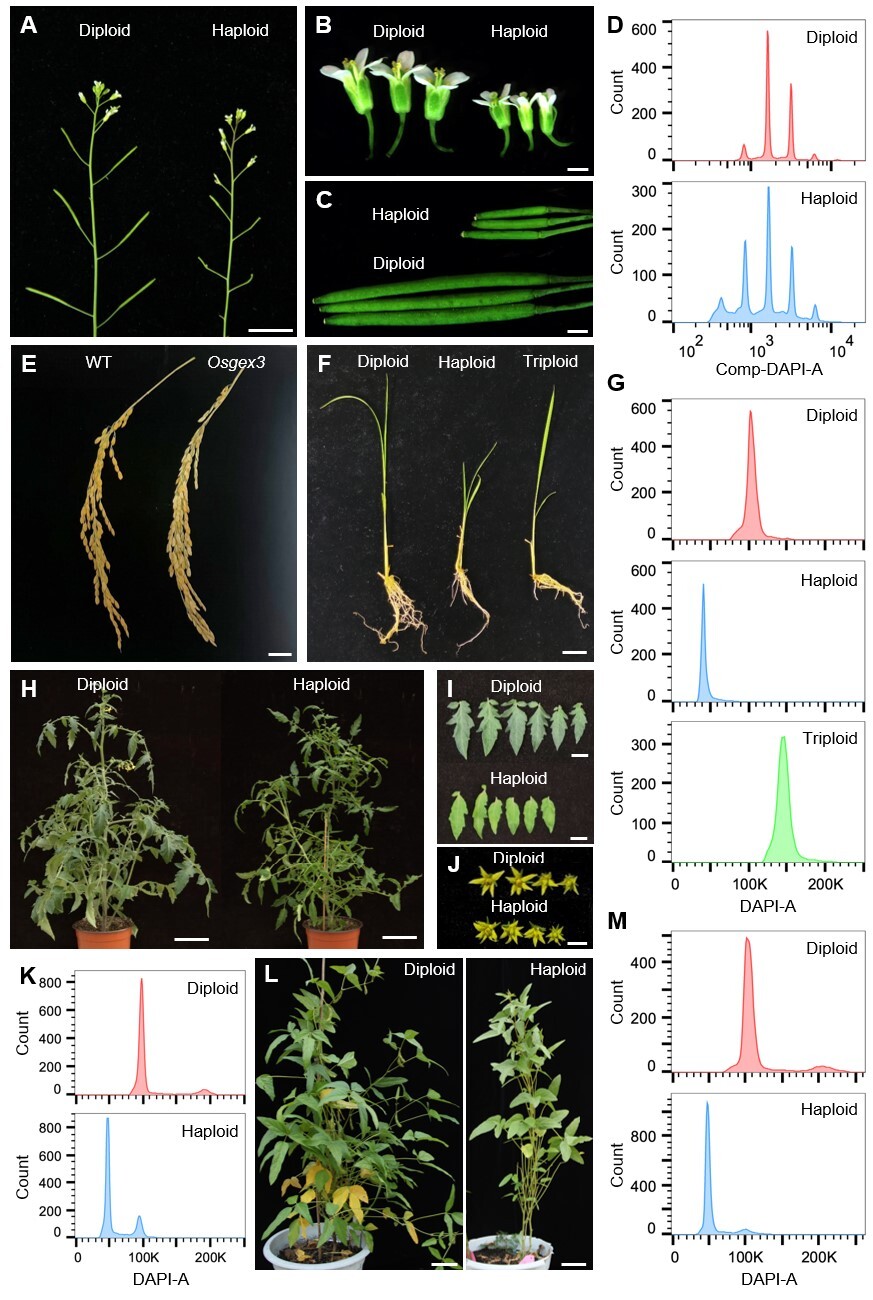
在第二项研究中,研究人员发现将GEX3突变后,在拟南芥gex3突变体中能鉴定到0.57%的单倍体植株,通过将DMP8/9同时突变,单倍体诱导率能提升至3.13%。研究人员还鉴定到GEX3在水稻、番茄和大豆中的同源基因,发现将它们突变后也能诱导单倍体产生,在大豆中的单倍体诱导率高达8.99%。值得注意的是,在水稻中,OsGEX3的缺失还能产生9.82%的三倍体后代,表明由GEX3调控的受精过程既具有保守性,又具有物种特异性差异。该研究为作物双单倍体技术和无融合生殖提供了新的基因资源。
GEX3突变诱导单倍体
这两项研究成果于2026年5月在线发表于
Plant Communications杂志(https://doi.org/10.1016/j.xplc.2026.101883、
https://doi.org/10.1016/j.xplc.2026.101884)。遗传发育所李红菊研究组博士后王岚为第一篇论文的第一作者,王岚和李红菊研究组助理研究员陈澍燕为第二篇论文的共同第一作者,遗传发育所李红菊研究员和杨维才院士为共同通讯作者。这两项工作得到了国家自然科学基金、国家重点研发计划、中国科学院战略性先导科技专项和中国科学院稳定支持基础研究青年团队项目的资助。